水分量の変化や気象状況の変化または測定条件が必ずしも実際の現象と一致しないことに起因する誤差と考えられる。しかし、各深さにおけるK1およびK2の平均値は、互いに比較的類似した分布となっている。すなわち、温度伝導率は表層付近で低く、逆に5cm〜10cmの深さで高くなり、さらにその下層ではそれらの平均的な値となっている。このため、潟土中の平均的な温度伝導率は4×10-3cm2/sec 程度と推測される。また、K1とK2とは1:1の直線の回りに散在しており、(4)式、(6)式いずれの式を用いてもほぼ同じような値が推算された。
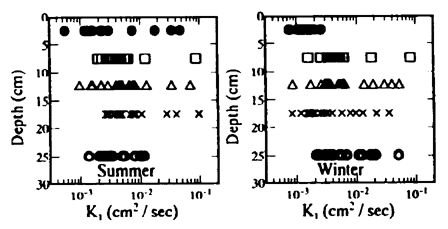
Fig.6 Relationship between K1 and tideland for sediment
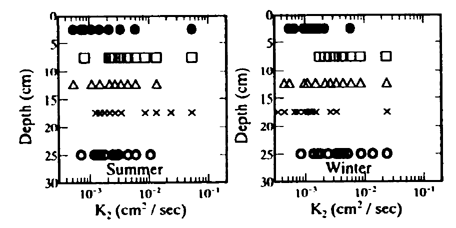
Fig.7 Relationship between K2 and tideland depth for sediment
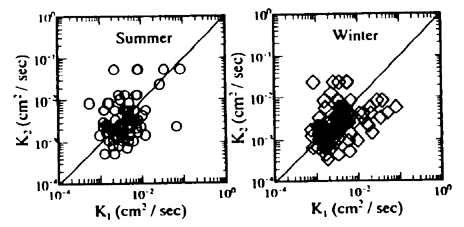
Fig.8 Relationship between K1 and K2 for sediment
3.4 基化連元電位
Fig.9は測定地点における酸化還元電位を示したものである。図示されるように有明海の干潟域の底層は、海水または表層土によって大気と遮断され、酸素の供給は制限される。そのため表層(酸化層)と下層(還元層)とは分化され異なる特徴を示す。酸化層から還元層にかわる酸化還元電位の不連続層(Redox Pottential Discontinuity Layer;RPD層)は季節ごとに若干の差はあるが、ほぼ2cm前後で形成されている。これは底泥の違いはあるもののN.P.Revsbechら6)の研究成果とほぼ同様な結果が得られた。
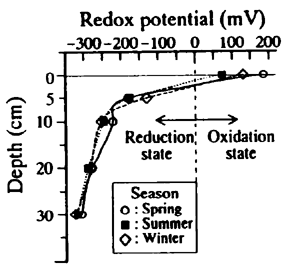
Fig.9 Seasonal profiles of the redox potential with depth
3.5 栄養塩濃度の変動特性
Fig.10は潟土間隙水中の栄養塩濃度の時間的および季節的変動特性を示した。なお、Fig.11は潟土間隙水中の栄養塩濃度別の空間的変動特性である。図示されるようにFig.9の酸化還元電位と同様酸化眉より還元層にかけて微生物群集のエネルギー代謝に関係する栄養塩の濃度を測定すると深さに応じて特徴的な鉛直分布を示す。すなわち、潟土中では酸化還元電位の鉛直的変化に対応して有機物分解に関与する細菌群のエネルギー代謝の形式が段階的に変化することを強く示唆している7)。干潟域の底質は表層の酸化層を除くとほぼ還元状態であるため、一般的な水域で見られるような硝化作用はほとんど見られず、窒素態は大部分アンモニア態窒素(NH+4-N)状態で存在する。すなわち、NH+4一Nの濃度変化は、2月を除く年間を通して高濃度で安定した濃度分布を呈する。これを気温分布と照らし合わせてみると、春・夏期には気温が上昇し生物の活動が盛んな時期で、有用生物からの排せつ物や死骸からのNH+4-Nの濃度も上昇し、間隙水中に存在する窒素態のほとんどはNH+4一Nとなっている。これは気温上昇にともない潟土中の嫌気性菌の活動で冬期に蓄積された硝酸態窒素(NO-3-N)や亜硝酸態窒素(NO-2-N)をNH+4-Nに変化させ、NH+4-N濃度を上昇させたものと考えられる。すなわち、NO-3-4の濃度分布は、5/15〜8/12の間で生物の活動に起因するNH+4-N濃度上昇に追いつかず、窒素態はほとんどNH+4-Nとして存在する。そして、気温の低下に伴い生物活動も弱まり細菌による硝化作用が生物活動に起因するNH+4-Nの濃度上昇に優り、NO-3-N濃度が上昇
前ページ 目次へ 次ページ